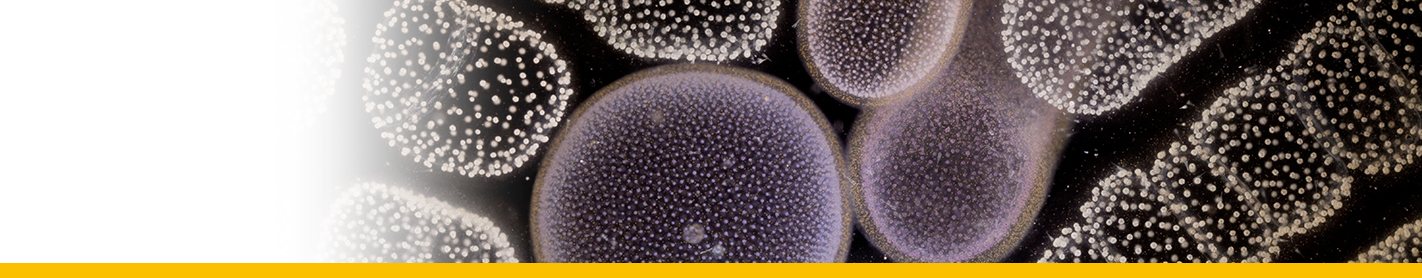
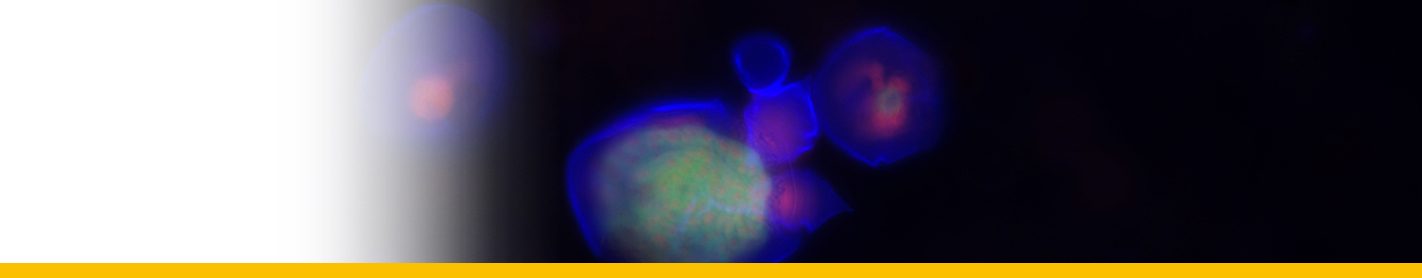
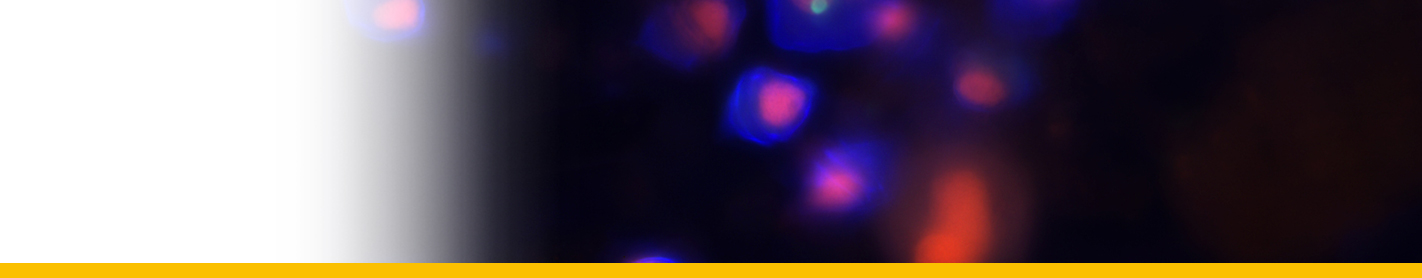
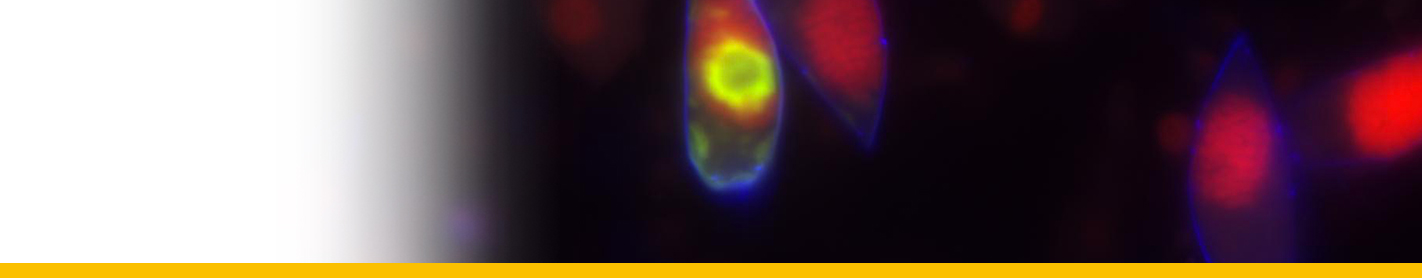


Euduboscquella crenulata
Diagnosis
Diagnosis_Genus: Euduboscquella Coats & Bachvaroff 2012. Euduboscquellidae with trophont episome as disc-shaped shield bordered by a perinematic ring. Lamina pharyngea extending from perinematic ring into trophont cytoplasm. Food vacuole formed as trophont emerges from host giving rise to extracellular tomont. Multiple spore morphotypes possible, including dinokont and non-dinokont cells. Individual infections producing only one type of spore.
Diagnosis_Species: Euduboscquella crenulata Coats & Bachvaroff 2012. Trophont variable in dimension depending on age; nucleus increasing in size with cell growth. Nucleolus present only in very young trophonts; nodes of heterochromatin distributed at periphery of nucleus in older trophonts. Surface of shield with numerous, irregular grooves giving crenulated appearance in optical section. Lamina pharyngea as a spiraled funnel with ~ 1.5 turns; 10--25 µm long. As the parasite emerged from the host cell, the shield curved inward and the perinematic ring closed, surrounding part of the host’s cytoplasm in a food vacuole. Sporogenesis starts outside the host. Three types of spores are produced: sigmoid dinokont spores (male spores) measuring 7--10 by 2--6 µm in vivo with subequatorial, ovoid nucleus; spherical spores (female spores) measuring 10--14 µm in vivo, with acentric, ovoid nucleus; egg-shaped (neuter) spores measuring 10--16 by 8--13 µm wide in vivo, with large vacuole filling most of cell; roughly spherical nucleus positioned at narrow end of cell. The nucleus of neuter spores had two or more nucleoli and sometimes appeared to have condensed chromosomes.
==> E. crenulata differs from closely related species in nuclear and cortical morphology of the vegetative trophont, including a distinctively grooved shield (= episome) that imparts a crenulated appearance in optical section.
Body_length: 10-25 µm
Body_spores_length: 7-10 µm
Body_spores_wide: 2-6 µm
Body_spores_length: 10-14 µm
Etymology
Genus name is derived from the Greek eu- (= well, normal) and the genus name Duboscquella.
Species name is from the Latin crenula meaning notch and is used in reference to the crenulated appearance of the shield in optical section.
Type species
This is the type species of the genus.
Type illustration / Type locality / Type specimen
Type host: Favella panamensis Kofoid & Campbell, 1929
Type habitat: Polyhaline tidal lagoon at the northern end of Ocean City, MD, USA.
Type locality: Assawoman Bay, Ocean City, MD, USA (38° 20’ 02” N; 75° 05’ 47” W)
Type material: An hapantotype slide of protargol-impregnated Favella panamensis infected by E. crenulata has been deposited in the International Protozoan Type Slide Collection, National Museum of Natural History, Washington, DC, USA under the registration number 1156917
Ecology
Substrate: endozoic (endoparasite)
Sociability: solitary
Salinity: marine
pH: neutral
Life cycle
E. crenulata produces three morphologically distinct spore types, two of which (female and male) conjugate to form a uninucleate zygote. The zygote undergoes successive division to produce four daughter cells of unequal size, but that resemble the non-mating spore type (the neutral spores). Coats et al. 2011.
More detailed: Male spores remained active for several hours, swimming erratically between short periods of quiescence, but never formed pairs suggestive of conjugation. A day or two after formation, male spores lost motility and degenerated. Female and neuter spores persisted for 2--3 days before degrading. Addition of fresh neuter spores to wells containing active male spores elicited no response. By contrast, addition of female spores generated an immediate reaction, with male spores forming a cloud of rapidly swimming cells around each spherical female spore. Only one male spore attached to the surface of each female, with the surrounding cloud of spores almost immediately dispersing. Prior to attachment, male spores developed a small protuberance or copulation globule on their ventral surface near the insertion of the flagella. Thus, conjugation occurred with the ventral surface and flagellar basal bodies of the male spore pressed against the female spore. The flagella of attached males remained active, causing the conjugating pairs to rotate rapidly. Motility soon ceased, as resorption of the male flagella accompanied fusion of the paired cells. Karyogamy preceded completion of cytogamy. Conjugation lasted 20-30 min, with the zygote remaining inactive for ~2 h before undergoing two successive divisions presumably representing meiosis. Daughter cells of the first division were of equal size and sometimes showed a spindle with metaphase arrangement of chromosomes as the second division proceeded. The second division was not equal, with the resulting chain of four cells having two small central cells flanked by larger individuals. The chain of post-meiotic daughter cells gradually dissociated, giving rise to small egg-shaped cells (7--11 µm by 5--8 µm in vivo) that resembled neuter spores. When 24 female spores were transferred to wells containing male spores, all were fertilized and underwent meiosis. The remaining population of female spores (44 total) retained in the absence of male spores failed to divide. Neuter spores did not divide whether maintained alone or incubated in the presence of male spores.
E. crenulata and E. cachoni each produce three types of spores: motile dinokont spores, non-motile spherical spores, and non-motile egg-shaped and spindle-shaped spores, respectively (Coats 1988; Coats et al. 1994). Coats (1988) suggested that the spindle-shaped spores of E. cachoni might be cysts, as they persisted in the lab for several days. By contrast, the egg-shapes spores of E. crenulata degenerated in 2-3 days when held at near ambient water temperature and are therefore not likely resting cysts.
Generation: <1 month
Reproduction_mode: sexual
Reproduction_mode: asexual
Symbiont: horizontal
Feeding behaviour
Mode of locomotion
Reference(s)
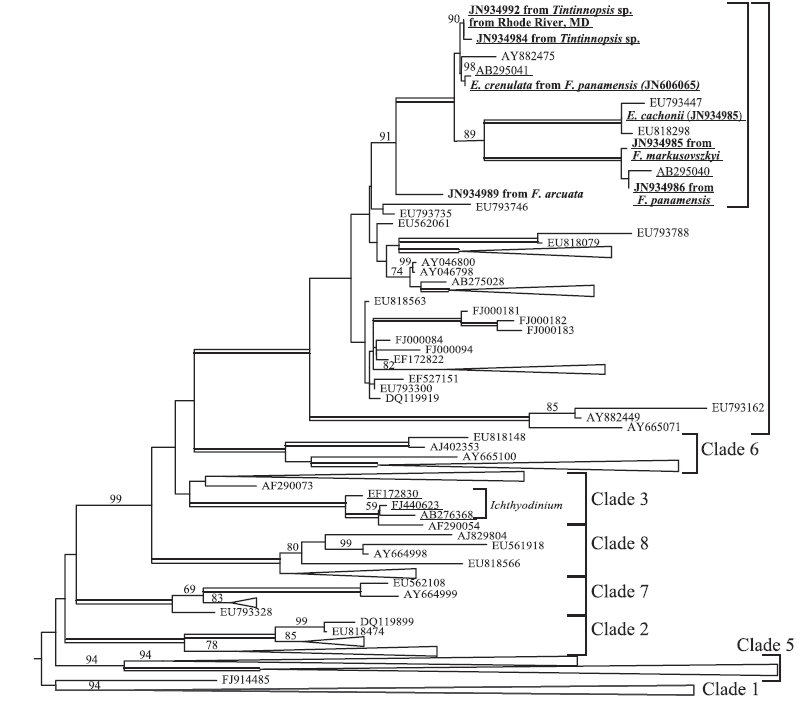
Attached phylogeny
Observation site(s)
HOSTS
| Association with... | Region origin | Name of site | In reference... |
|---|---|---|---|
| Favella ehrenbergii | Shizuoka | Hamana Lake |
(2007) Species of the Parasitic Genus Duboscquella are Members of the Enigmatic Marine Alveolate Group I. Protist 158:337 - 347. doi: 10.1016/j.protis.2007.03.005 |
| Favella panamensis | Maryland | Assawoman Bay | |
| Favella panamensis | Assawoman Bay |
(2012) Molecular Diversity of the Syndinean Genus Euduboscquella Based on Single-Cell PCR Analysis. Applied and Environmental Microbiology 78:334 - 345. doi: 10.1128/AEM.06678-11 |
|
| Favella panamensis | York River |
(2012) Molecular Diversity of the Syndinean Genus Euduboscquella Based on Single-Cell PCR Analysis. Applied and Environmental Microbiology 78:334 - 345. doi: 10.1128/AEM.06678-11 |
|
| Favella panamensis | Florida | Indian River |
(2012) Molecular Diversity of the Syndinean Genus Euduboscquella Based on Single-Cell PCR Analysis. Applied and Environmental Microbiology 78:334 - 345. doi: 10.1128/AEM.06678-11 |
| Favella panamensis | Maryland | Solomons Island |
(2012) Molecular Diversity of the Syndinean Genus Euduboscquella Based on Single-Cell PCR Analysis. Applied and Environmental Microbiology 78:334 - 345. doi: 10.1128/AEM.06678-11 |

{kind=link}